
玉米产量相关性状的遗传解剖和这些性状的中亲优势


文|名城雨
编辑|名城雨
利用玉米优势在玉米育种中对提高籽粒产量至关重要。然而,优势的遗传结构尚不完全清楚。为剖析玉米产量相关性状和优势的遗传基础,选取了301-08个×YE641的478个重组自交系和永生化F的298个杂交系2群体用于绘制9个产量相关性状和中亲优势的数量性状位点(QTL)。
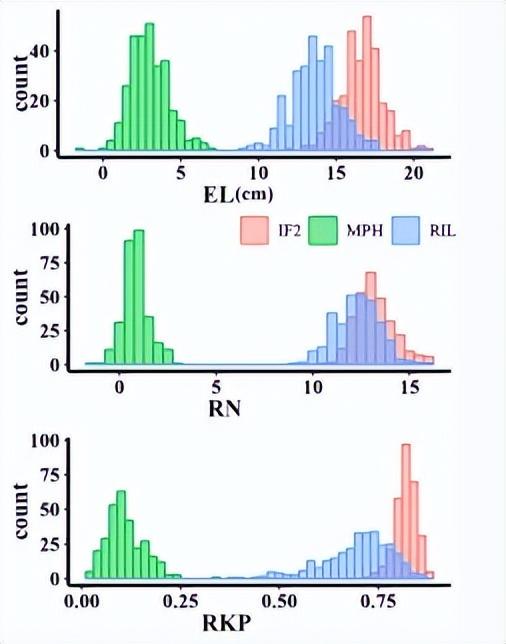
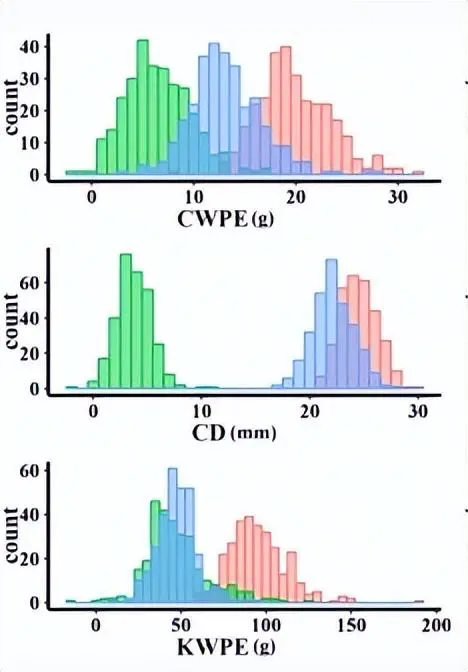
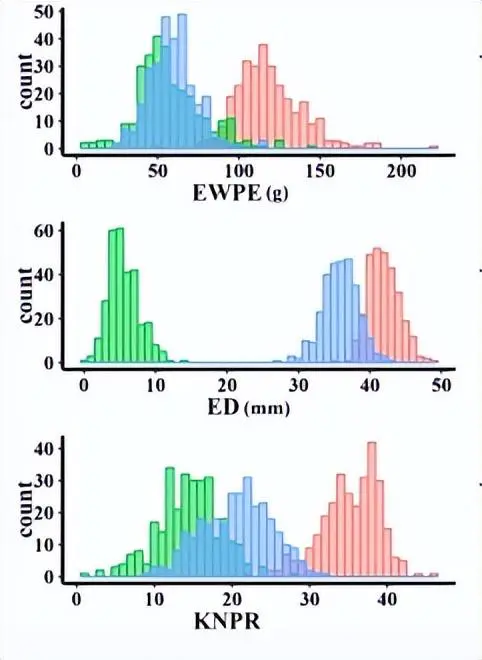
我们在近交和杂交映射群体中观察到156个QTL,28对具有上位相互作用的位点和10个显着的QTL×环境相互作用。F中的高优势1和 IF2对于表现出优势的性状,每耳的内核重 (KWPE)、每耳的耳重 (EWPE) 和每行内核数 (KNPR) 的种群与表现出优势的性状的高百分比 QTL (超过 50%)相匹配,而对于表现出低优势的性状,例如每穗玉米芯重 (CWPE), 果仁产量 (RKP)、穗长 (EL)、穗径 (ED)、玉米芯直径和行数 (RN)。
环境稳定的QTL qRKP70-3在两个映射群体中被鉴定出来,而qKWPE2影响性状均值和中亲优势(MPH)水平,解释了超过9%的表型变异。18个QTL,qEWPE9-1,qEWPE10-1,qCWPE6,qEL8,qED2-2,qRN10-1,qKWPE9,qKWPE10-1和qRKP4-3,占表型变异的10%以上。 此外,QTL 映射确定了 95 个 QTL,这些 QTL 被收集在一起并整合到 33 条染色体上的 10 个 QTL 簇中。
结果表明:(1)异质模式改善Reid(PA)×Tem-tropic I(PB)中产量相关性状和MPH的遗传具有依赖性;(2)很大一部分位点表现出优势效应,而优势也对KNPR、EWPE和KWPE的MPH;(3)在基因组区域1.09-1.11、2.04、3.08-3.09和10.04-10.05的标记辅助选择有助于杂交性能本身和优势,并且在以前的研究中反复报告使用不同的模式是推荐的。
利用玉米优势对提高籽粒产量具有重要意义。到二十世纪后期,玉米杂交种占玉米总种植量的65%,并使玉米年产量翻了两番。许多育种者多年来一直对优势感兴趣;然而,对与优势有关的遗传学对谷物产量的理解仍然难以捉摸。因此,研究和评估玉米优势对籽粒产量的遗传机制,将为理解这些现象铺平道路,有助于优化不同群的产量育种。
通过早期的近交选择和实验育种,玉米育种者认识到种质资源和优势的重要性。以前的研究使用分子标记,表型鉴定和谱系信息将不同的种质分为不同的群。异质模式硬茎合成(SS)×非硬茎(NSS)已被广泛用于玉米育种。
Li和Wang在以前的研究基础上,提出了五个群,即唐思平头(TSPT),兰开斯特,兰开斯特样,改进的Reid(PA)和Tem-tropic I(PB)。TSPT×Reid杂交模式得到了广泛的研究,因为它是中国北方主要的模式之一,并已用于生产正单958等精英杂交种。
然而,这种异性模式在中国西南部很少使用,在那里,PB×PA等替代的、很少研究的模式更合适。因此,本研究研究了PB×PA杂交种中产量相关性状的优势遗传学以及产量相关性状本身的遗传学。
本研究旨在剖析玉米籽粒产量相关性状的遗传结构以及不同遗传效应对籽粒产量相关性状优势的贡献,本文在301种环境下评估了08-641×YE478的多个RIL和IF。在三种环境下,这些RIL产生的种群。精英系YE478,属于PA杂交组,是中国杂交基础之一。
另一亲本08-641(SAU08-641)来源于PB(或Tem-tropic I杂交群)种质,是自1998年以来在中国西南地区广泛使用的典型优良自交系。PA×PB是中国西南地区主要的模式之一。本研究的主要目的是 (1) 使用 RIL 群体检测和估计对产量相关性状具有加性和加性加性效应的 QTL (2) 使用源自 RIL 群体的杂交群体 (IF2种群)(3)使用映射种群鉴定中亲优势(MPH)的QTL,以及(4)确定与谷物产量和MPH相关的最合适的基因组区域,通过标记辅助选择进行管理。
在RIL和IF方差的组合分析中,在所有环境中,大多数产量相关性状的基因型、环境和基因型×环境相互作用的变异来源都非常显著(P < 0.01)。广义遗传力(h2)的性状范围从KWPE的60.8%到RN的91.1%。EWPE和KWPE的遗传力在IF中人口比 RIL 人口更大。
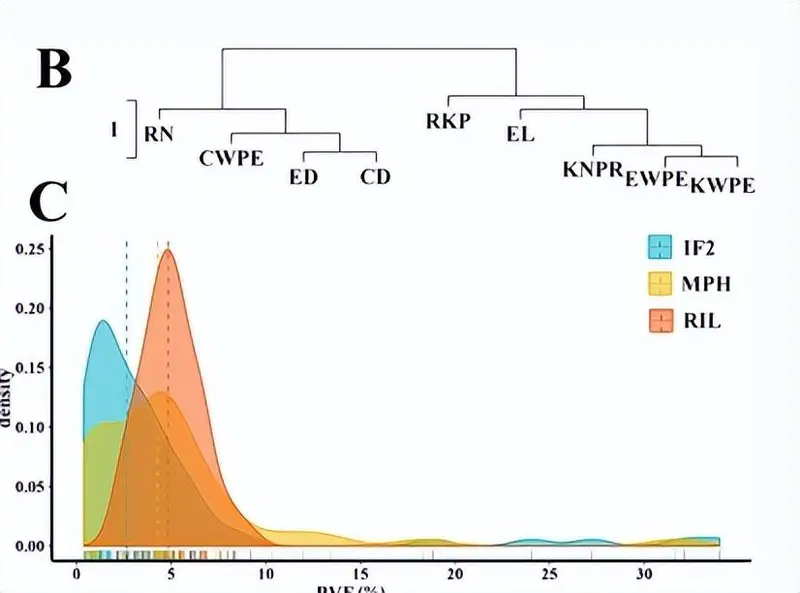
产量相关性状与这些性状的中亲优势(MPH)之间的相关性关系,以及不同数据集中的QTL分布。a 产量相关性状与MPH之间的皮尔逊相关系数(|r2|> = 0.5)。黄线表示在 MPH 数据集中发现的显著相关性;蓝线表示RIL的显著相关性;绿线表示两个IF的显著相关性2和 MPH 数据集;黑线表示在RIL中发现的显着相关性,即IF。2和 MPH 数据集。
在目前的研究中,在所有环境中发现了156个QTL的平均产量相关性状和这些性状的MPH, 通过单一环境分析,在至少一个环境中还鉴定了跨环境检测到的 6 个 QTL。这些QTL分布在所有染色体上,尽管在bin 08.3-09.10和04.10-05.0处发现了两个QTL浓缩区域。
每个QTL解释了38.33%至99.5%的表型方差(PVE),超过一半的QTL各占PVE的不到2%,QTL数量和总PVE在性状和种群之间差异很大。在RIL和IF中同时检测到多个QTL2群体,而在 IF 中检测到 13 个 QTL2人口和在MPH数据集中共同定位。
在对 MPH 进行 QTL 分析时,在 RIL 群体中发现的只有两个 QTL 也位于同一区域。这些结果表明,参与杂交性能本身的遗传学可能部分与MPH相关,并且与IF中杂交性能本身之间的中高相关系数一致。除RKP外,所有性状都报告了总体和相应的MPH水平。
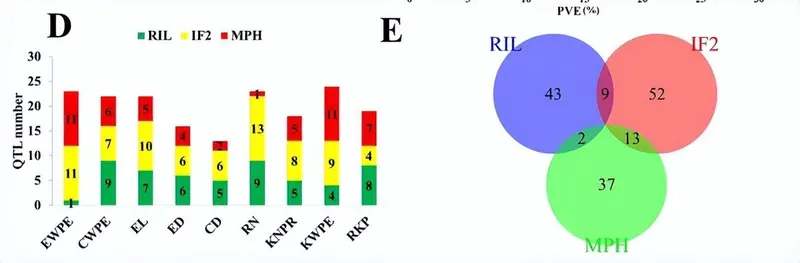
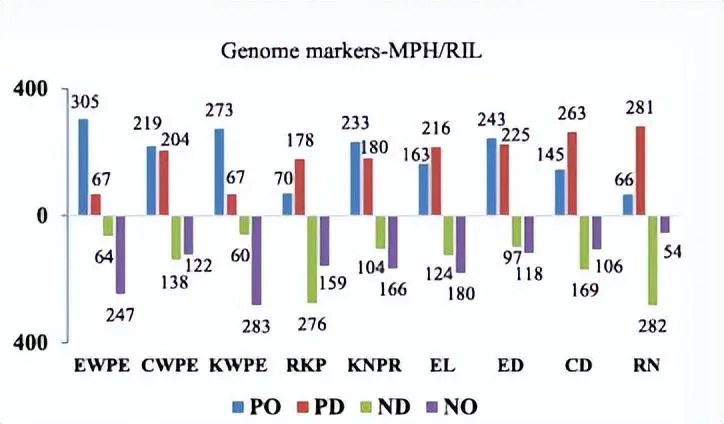
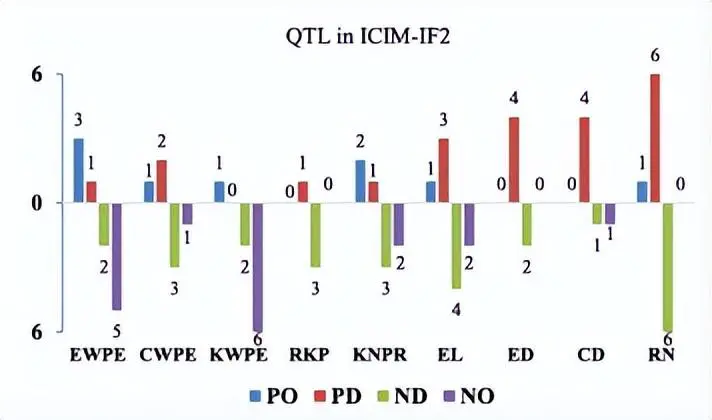
在RIL和IF中检测到的产量相关性状的QTL分布2种群和优势。红色矩形表示具有负面影响的 QTL(RIL 群体中的加性,IF 中的显性和加性)2中亲优势(MPH)的种群和优势效应),而蓝色矩形表示积极影响。矩形的宽度表示 QTL 的置信区间。十条染色体的QTL密度热图显示在图的底部和X轴下方。EWPE,每只耳朵的耳朵重量;CWPE,每只耳朵的玉米芯重量;EL,耳朵长度;ED,耳径;CD,玉米芯直径;RN,行号;KNPR,每行内核数;KWPE,每耳果仁重量;RKP,内核生产率。
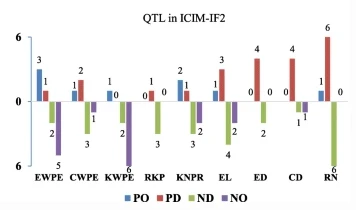
我们使用ICIM-EPI在全基因组搜索中通过上位位点估计了总PVE,在 IF 中2人口,上位点在CWPE(50% vs 41%)、ED(24% vs 20%)、CD(56% vs 32%)和KNPR(53% vs 37%)方面的PVE高于单位点QTL。在MPH数据集中,除EWPE(20%对49%)、CWPE(23%对28%)、KWPE(43%对52%)和RKP(16%对43%)外,所有性状的上位点总PVE均大于单位点QTL。
表明EPI对这些性状的影响大于单位点QTL,无论统计学意义如何。目前的研究结果还表明,结合上位点和单位点QTL产生的累积效应解释了PVE的很大一部分。同时,基于混合模型的复合区间映射(MCIM)和全QTL模型,28对标记区间表现出显著的上位相互作用效应;在RIL群体中检测到13个加法(AA)相互作用,在IF群体中检测到10个上位上位相互作用2种群和 MPH 数据集中的五个二基因相互作用。
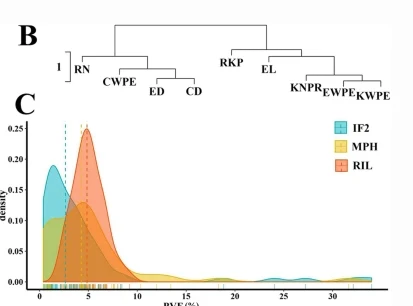
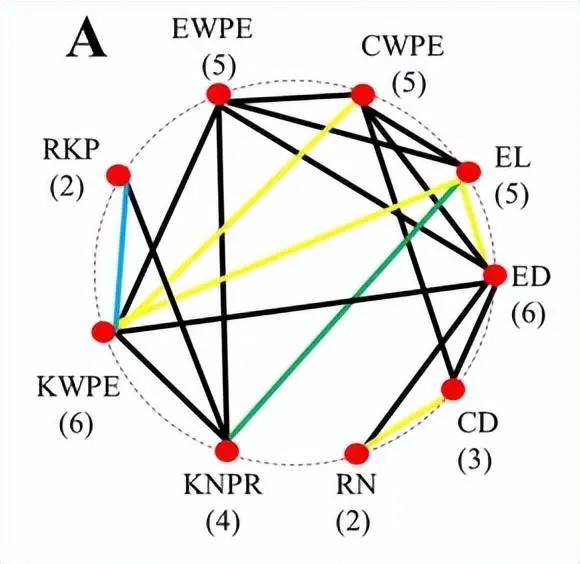
不同性状的重叠 QTL 被分配到同一个 QTL 集群。总共有95个QTL被分组为分布在所有染色体上的33个QTL簇。每个 QTL 集群包含 3 到 3 个特征的 QTL。包含六个QTL的簇QC10-3影响了EL,RN,ED,KNPR,RKP和KWPE的平均值,以及RKP的MPH。QC5-10在60号染色体的1 cM区域内,对EWPE,KNPR,CWPE以及ED和CD的MPH水平具有多效性作用。
正如杂交性能本身与MPH之间中度至高度相关性所预期的那样,与产量相关性状和MPH相关的一些位点聚集在一起。近5%的QTL簇与两个性状相关。其中一些聚类是针对确定的人群或MPH的特异性。
本研究中使用的绘图种群由301个RIL组成,这些RIL来自玉米自交系08-641和Ye478之间的杂交。精英自交08–641来自四川农业大学玉米研究所,Ye478由莱州市农业科学院制定的国家重大基础乳品研究“973”计划项目提供。
在每个样地中,从所有收获的耳朵中随机选择10个授粉良好的耳朵。测量的九个产量相关性状是:EL(厘米),ED(毫米),CD(毫米),RN(计数),KNPR,EWPE(g),CWPE(g),KWPE(g)和RKP。每次复制的均值用于进一步分析。使用统计软件包SPSS 17.0中的PROC CORR进行表型性状的分布和皮尔逊相关性的分布,使用IF的301个RIL和298个杂交种中的表型性状进行2所有环境中的人口。
基于RIL的标准化数据,对表型性状进行了分层聚类分析(“hclust”)。RIL和IF方差的组合分析2使用SPSS 17.0中的GLM程序计算每个性状的群体,基因型为固定效应,复制和环境为随机效应。广义遗传力(h2) 用于 RIL 和 IF2人口是在Hallauer和Miranda描述的条目平均值基础上计算的。
综合分析,估计种群的产量相关性状。对于一维基因组扫描,测试窗口和过滤窗口尺寸设置为10 cM,步行速度设置为2 cM。在1000个排列检验的帮助下估计F统计量和临界F值,p<0.05作为候选区间选择的实验显著性水平。
每个性状的最终完整QTL模型包含显着的加法,优势和上位效应,以及它们与环境的相互作用。假设实验误差<0.05,则使用Bonferroni校正来计算比较显著性阈值。我们还计算了每个RIL的杂合位点百分比,分析了表型性能与杂合性之间的相关性关系,并对杂合性上的表型性能进行了回归分析。
参考文献:
- 优势:养活人们和保护自然资源。在:库尔斯JG,潘迪S,编辑。作物优势遗传学和利用国际研讨会论文集,CIMMYT,墨西哥城,麦迪逊
- 商业杂交玉米育种计划的长期选择。在:Janick J,编辑。植物育种评论24,第2部分。霍博肯:威利;2004.第109–51页。
- 杜维克·育种对玉米产量进步的贡献。阿格隆。2005;86:83–145.
- 慈鑫, 李敏, 梁晓, 谢志, 张丹, 李鑫, 等.1970—2000年中国玉米杂交种对高产的遗传贡献.作物科学. 2011;51:13–20.
- 莫尔,隆奎斯特,贝莱斯,约翰逊·玉米优势与遗传分化的关系.遗传学。1965;52:139–44.
- Moll RH, Salhuana WS, Robinson HF. 玉米品种杂交的优势和遗传多样性.作物科学. 1962;2:197–8.
- Smith OS, Smith JS, Bowen SL, Tenborg RA, Wall SJ.一组优良玉米自交系之间的相似性,通过血统、F1粒产量、籽粒产量、优势和 RFLP 来衡量。1990;80:833–40.
- BeckDL, Vasal SK, Crossa J. 亚热带和温带中熟玉米种质的优势和结合能力.作物科学. 1991;31:68–73.
- Messmer MM, Melchinger AE, Boppenmaier J, Herrmann RG, Brunklaus-Jung E. 早熟欧洲玉米种质的RFLP分析。I. 燧石和凹痕近交系的遗传多样性。Theor Appl Genet。1992;83:1003–12.
- Messmer MM, Melchinger AE, Herrmann R, Boppenmaier J. 早熟欧洲玉米杂交种之间的关系.作物科学II 1993;33:944–50.